
Integrin Mediated Mechanotransduction
Integrins are heterodymeric proteins composed of a and ß subunits responsible for adhesion of cells to the extracellular matrix (ECM) and are very involved in a host of signaling pathways. There exists a wide variety of specific a and ß subunits, with each combination binding to specific amino acid sequences present in the ECM. Commonly expressed integrins include a5ß1 and avß3,which bind to amino acid sequencs in fibronectin and vitronectin respectively. Structurally, the a subunit is composed of two polypeptide chains linked via a disulfide linkage. The ß subunit on the other hand is composed of a single polypeptide chain. The entire heterodymeric complex is itself complexed to divalent cations, without which binding ability is severly compromised. Integrins provide a direct connection between the actin cytoskeleton and ECM forming focal adhesions. Integins are bound to the actin cytoskeleton through at least four known proteins, one of the most important being Talin, which plays an early role in reinforcing focal adhesions under stress. Integrins play crucial roles in many “outside-in” and “inside-out” signaling cascades. Signaling cascades in general are very complex and not completely understood, but here we shall introduce a few of the players. Focal adhesion kinase (FAK) is a protein tryrosine kinase activated by integrin ligation and has been shown to play a vital role in mechanosensing. Rac is a downstream GTPas
e target of FAK that plays a role in reoganization of the actin cytoskeleton into an isotropic network. Rho another small GTPase is downregulated upon FAK activation and is responsible for organizing the actin cytoskeleton into stress fibers. Rho and rac behave antagonistically with regard to actin organization. A schematic of this cascade is shown here.1
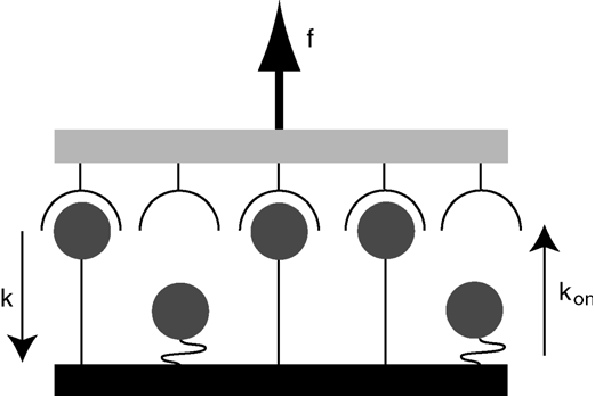
It is instructive to consider a parallel binding model (shown left) in the case of focal adhesion mechanotransduction. A collection of integrins are bound to the ECM in a parallel manner, such that when a force is applied to the surface of the cell, the said force is shared equally between each integrin attachment. This is the case when the ECM is considered a soft spring, in the opposite limit of a stiff spring cooperativity is lost. Integrin-ECM associations are assumed to rupture as a function of applied force with a rate of k = k0eF/iFb, where k0 is the unstressed dissociation rate, F is the applied force and Fb is the force scale (typically pN). The factor i is introduced as a result of cooperativity. Once dissociated, the bonds can reform at a rate of kon. This leads to:
Where N is the mean number of bound receptors, t is dimensionless time (t=k0t), f is dimensionless force (F/Fb), ? is dimensionless rebinding rate (?=kon/k0) and Nt is the cluster size. This equation essentially states that the rate of change in the number of closed bonds is given by the difference of the number of bonds forming and the number of bonds rupturing due to an applied force. Applied forces destablize the cluster, which is restablized by rebinding. By setting dN/dt=0 we can find the critical force fc, below which there is net binding and above which there net dissociation:
Where plog(a) is defined as the solution of xex=a. This implies that focal adhesions regulated close to fc would provide fine control in dynamic mechanical loading, as increased forces would result in a net loss of integrin signaling and decreased forces would result in a net gain.1
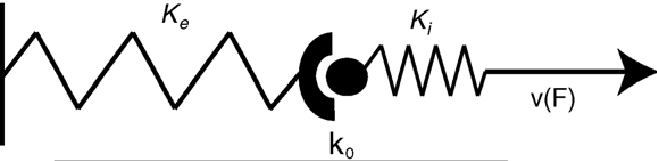
This model can be extended by viewing the actin-integrin complex as elastic along with the ECM ligands (shown above), each with an associated spring constant ki and ke respectively. A receptor ligand complex could then be represented by two springs in series. Tension is generated in the actin stress fibers via the molecular motor myosin, thus ki is much stiffer than ke, and the complex has roughly the stiffness of the softer spring (1/k = 1/ki + 1/ke), in this case the ECM. The force generated by the actin-myosin cross-bridge can be modeled using a linearized force velocity relationship:
Where v0~10 µm/s and Fs is the stall force. As the myosin-actin-integrin complex pulls on the ECM, elastic energy is strored in the receptor ligand complex W=F2/2k. Therefore, the stiffer the spring complex, (ECM) the less work must be done to mechanical store energy in the receptor-ligand complex. Taking the derrivative with respect to time gives the relationship for power, using the linearized force velocity eqaution above gives:
This can be integrated to yield:
Thus, individual receptor-ligand complexes can generate pN forces on millisecond time scales (typical time scale is tk) given typical parameters. Thus, individual receptor-ligand complexes and focal adhesions on the whole can be modeled and the force required to rupture is given by:1
Thus, as shear stress is increased, there is an increasing force applied to the focal adhesions required to keep the endothelial cell from being swept away. This influences integrin-ECM binding in the manner described above, leading to the signaling cascade, which regulates cell function.
--------------------------------------------------------------------------------------------------------------
1. Ulrich S. Schwarz, Thorsten Erdmann and Ilka B. Bischofs. "Focal ashesions as mechanosensors: The two-spring model." BioSystems 83 (2006) 225-232
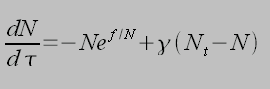
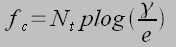
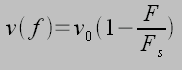
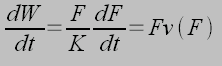
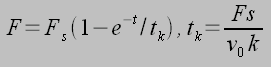
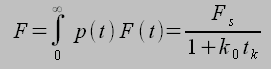
This page was last edited 10 June 2008